 Boka: 'Adam and the Genome' og "Ikke-funnksjonelle" pseudogener
Boka: 'Adam and the Genome' og "Ikke-funnksjonelle" pseudogener
Evolution news; 24. jan. 2018
Oversatt herfra.
Bilde 1. Høne; kredit: pexels, via Pixabay.
I (boka) 'Adam and the Genome' , som vi har vurdert i denne serien , hevder den teistisk evolusjonistiske biologen Dennis Venema at et annet bevis som felles design ikke kan forklare, er tilstedeværelsen av delte pseudogener hos mennesker og andre organismer, som aper. Ved første øyekast virker det som om han har et poeng: Mens felles design enkelt forklarer funksjonelle likheter, er ikke-funksjonelle likheter trolig best forklart av blindmekanistiske prosesser som mutasjon.
Pseudogener er relevante for spørsmålet om menneske/ape felles stamfedre, men hvis du definerer Adam og Eva som et første par som var historiske mennesker og forfedre for hele menneskeheten, kan du potensielt akseptere felles forfedre, men tro på en tradisjonell Adam og Eva. Som sådan er pseudogener ikke strengt tatt relevante for Adam og Eva som menneskets foreldre.
 Åpenbart, noen Bibel-troende som holder på en tradisjonell, historisk Adam og Eva, avviser felles forfedre. Men det er mulig å akseptere et historisk første par som menneskehetens forfedre, og likevel tro på felles avstamning. Noen forskere assosiert med BioLogos har fremmet denne ideen, med Adam og Eva som medlemmer av en stor befolkning av mennesker som samlet var forfedre for mennesker. Under denne oppfatningen er de ikke menneskets eneste forfedre, men de er forfedre for alle levende mennesker, sammen med andre. David Klinghoffer diskuterte denne oppfatningen her. Han bemerker at det holder seg til en historisk Adam, "men ikke i den forstand at de fleste kristne (og jøder og andre som leser Bibelen) forstår - et første par som menneskets eneste forfedre." I denne forstand er pseudogener Ikke nødvendigvis relevant for hvorvidt Adam og Eva eksisterte.
Åpenbart, noen Bibel-troende som holder på en tradisjonell, historisk Adam og Eva, avviser felles forfedre. Men det er mulig å akseptere et historisk første par som menneskehetens forfedre, og likevel tro på felles avstamning. Noen forskere assosiert med BioLogos har fremmet denne ideen, med Adam og Eva som medlemmer av en stor befolkning av mennesker som samlet var forfedre for mennesker. Under denne oppfatningen er de ikke menneskets eneste forfedre, men de er forfedre for alle levende mennesker, sammen med andre. David Klinghoffer diskuterte denne oppfatningen her. Han bemerker at det holder seg til en historisk Adam, "men ikke i den forstand at de fleste kristne (og jøder og andre som leser Bibelen) forstår - et første par som menneskets eneste forfedre." I denne forstand er pseudogener Ikke nødvendigvis relevant for hvorvidt Adam og Eva eksisterte.
Bilde 2. Kjente funksjoner hos 'junk-DNA'
Når det er sagt, ville delte pseudogener være best forklart av felles forfedre hvis (a) pseudogener er virkelig ikke-funksjonelle og (b) andre forklaringer som hotspot-mutasjoner ikke kan forklare disse delte likhetene.
Venema påstår at pseudogener er "ikke-funksjonelle". (S. 34) Problemet med hans argument fra pseudogener er at det nå er gode bevis på at pseudogener kan ha funksjon. Pseudogener kan produsere funksjonelle proteiner, funksjonelle RNA-transkripsjoner, og t.o.m. ha funksjon hvis de ikke produserer en transkripsjon. Følgende artikler har rapportert funksjoner for pseudogener:
Zheng and M. B. Gerstein, "The ambiguous boundary between genes and pseudogenes: the dead rise up, or do they?," Trends in Genetics 23 (May, 2007): 219-224
Hirotsune et al., "An expressed pseudogene regulates the messenger-RNA stability of its homologous coding gene," Nature 423 (May 1, 2003): 91-96
H. Tam et al., "Pseudogene-derived small interfering RNAs regulate gene expression," Nature 453 (May 22, 2008): 534-8
Pain et al., "Multiple Retropseudogenes from Pluripotent Cell-specific Gene Expression Indicates a Potential Signature for Novel Gene Identification," Journal of Biological Chemistry 280 (February 25, 2005):6265-6268
Zhang et al., "NANOGP8 is a retrogene expressed in cancers," FEBS Journal 273 (2006): 1723-1730
Faktisk rapporterte ENCODE-prosjektet over 850 humane pseudogener som er "transkribert og forbundet med aktivt kromatin."
Selvfølgelig er det mange som antas å være "pseudogener" i primat-genomene som vi ennå ikke har identifisert noen funksjon for. Venema identifiserer syv andre pseudogener relatert til olfaktoriske reseptorer som han hevder er "ikke-funksjonelle". Og det er mange andre. Problemet med Venema er at forskere som studerer pseudogener, er forsiktige mot å antage at pseudogener er ikke-funksjonelle, siden vi bare begynner å forstå dem.
En 2012-artikkel i 'Science Signaling' bemerket at selv om "pseudogener lenge er blitt avvist som søppel-DNA", har nyere fremskritt fastslått at "DNA fra et pseudogen, RNA transkribert fra en pseudogen, eller proteinet oversatt fra et pseudogen kan ha flere, mangfoldige funksjoner, og at disse funksjonene kan påvirke ikke bare deres opphavelige-gener, men også ikke-relaterte gener. " Artikkelen konkluderer med at" pseudogener har oppstått som en tidligere ukjent klasse av sofistikerte modulatorer for genuttrykk. "(Laura Poliseno, "Pseudogenes: Newly Discovered Players in Human Cancer," Science Signaling 5 (242) (September 18, 2012).)
En 2011-artikkel i tidsskriftet RNA er i overenstemmelse med dette:
 Pseudogener har lenge vært merket som "junk" DNA, mislykkede kopier av gener som oppstår under utviklingen av genomene. Men de siste resultatene utfordrer denne oppnavnet; Faktisk synes noen pseudogener å ha potensiale til å regulere sine proteinkodende fettere.
Pseudogener har lenge vært merket som "junk" DNA, mislykkede kopier av gener som oppstår under utviklingen av genomene. Men de siste resultatene utfordrer denne oppnavnet; Faktisk synes noen pseudogener å ha potensiale til å regulere sine proteinkodende fettere.
(RC Pink et al., "Pseudogenes: Pseudo-functional or key regulators in health and disease?" RNA 17 (2011): 792-798)
Bilde 3. Mot 'junk-DNA'
Fordi et pseudogen bare kan fungere i spesifikt vev og / eller bare i bestemte utviklingsstadier, kan deres sanne funksjoner være vanskelig å oppdage. Men en artikkel fra 2012 i RNA-biologi bemerker at "pseudogener ble lenge betraktet som søppel genomisk DNA", men "pseudogen regulering er utbredt i eukaryoter." Som det konkluderes med at "studien av funksjonelle pseudogener er bare i begynnelsen" og forutsier "mer og mer funksjonelle pseudogener vil bli oppdaget når nye biologiske teknologier utvikles i fremtiden. "(Wen et al.," Pseudogenes er ikke pseudo lenger ", RNA Biology , Vol. 9 (januar 2012): 27-32.)
Viktigst, når vi studerer nøye pseudogener, finner vi ofte funksjon. En artikkel i Årlig gjennomgang av genetikk observert: "Pseudogener som har blitt undersøkt, viser ofte funksjonelle roller." (Balakirev og Ayala, "Pseudogenes: Are they junk" eller functional DNA ?, " Annual Review of Genetics , Vol. 37: 123-51 (2003).)
 Venema antar at pseudogenene han undersøker er ikke-funksjonelle. Men det er gode grunner til å stille spørsmål om denne antakelsen. Det er mulig at i fremtiden vil mange flere funksjoner bli oppdaget.
Venema antar at pseudogenene han undersøker er ikke-funksjonelle. Men det er gode grunner til å stille spørsmål om denne antakelsen. Det er mulig at i fremtiden vil mange flere funksjoner bli oppdaget.
Som et kjent eksempel, under Kitzmiller v. Dover- rettssaken 2005, vitnet Kenneth Miller at den humane beta-globin-pseudogen er "ødelagt" fordi "den har en rekke molekylære feil som gjør genet dys-funksjonelt." Siden mennesker, sjimpanser , og gorillaer deler "matchende feil" i pseudogenet, vitnet han, "fører dette oss bare til en konklusjon ... at disse tre artene deler en felles stamfar." Men en 2013-studie i 'Genome Biology and Evolution' fant imidlertid at beta-globin pseudogen er funksjonell på grunn av sin "konserverte" sekvens som antyder en valgbar funksjon, noe som gjør den mindre tolerant for mutasjoner.
Bilde 4. Funksjonelle former av proteiner
For detaljer, se:
* "Dover Revisited: Med Beta-Globin Pseudogen nå funnet å være funksjonell, biter et ikon av "Junk DNA" -argument i støvet "-her.
*Ana Moleirinho et al., "Evolutionary Constraints in the ?-Globin Cluster: The Signature of Purifying Selection at the ?-Globin (HBD) Locus and Its Role in Developmental Gene Regulation," Genome Biology and Evolution 5 (2013): 559-571)
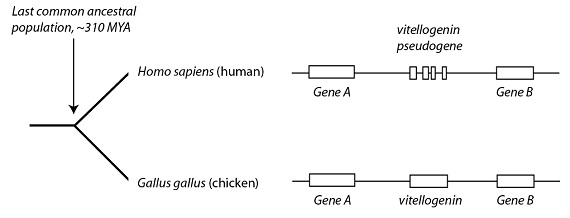
Som et annet eksempel, nevner Venema vitellogenin pseudogenet som angivelig demonstrerer felles forfedre mellom mennesker og fugler, som kyllingen. Kyllinger har tre vitellogenin gener som bidrar til å produsere eggeplomme. Mennesker legger åpenbart ikke egg, så antagelig trenger vi ikke gener til å produsere eggeplomme. Venema hevder imidlertid at mennesker har rester av tre ikke-funksjonelle pseudogener av vitellogenin (VIT1, VIT2 og VIG3) som vi arvet fra et eggleggende reptil -felles forfedre som vi deler med kyllinger.
Dette er ikke tilfelle hvor mennesker har en klar, fullstendig (eller til og med delvis fullført) ikke-funksjonelt pseudogen som tydelig samsvarer med et funksjonelt gen i noen beslektede arter. Snarere, ved Venema egen innrømmelse, har vi det han kaller "pseudogene fragmenter." Og med "fragmenter" snakker vi ikke om store DNA-strekk som tydeligvis samsvarer med store deler av leng-tapte vitellogenin-gener som fortsatt er beholdt hos våre kyllingfettere. I stedet merker han at disse er i beste fall "veldig små" og "små fragmenter av sekvens." (S. 39) Og viktigere, vi vil se nedenfor, Ann Gauger gjennomgikk disse justerte sekvensene og fant at de er så små at de er "på grensen av det som kan oppdages som en match", og konkluderer med at "matchen er svak".
Ikke desto mindre er Venema imponert fordi "disse små fragmentene er i riktig rekkefølge og kan stilles opp mot kylling-regionen." (S. 40) Han hevder at dette "rikelige" evidensbeviset betyr at folk bare avviser felles forfedre "på grunn av tidligere religiøse forpliktelser, ikke av vitenskapelige grunner."(s. 40) Men er det vitenskapelige beviset så entydig klart? Vel, la oss undersøke hans henvisning til disse påstandene.
Bilde 5. Villogenine -psedogen hos mennesket?
 Venema sitering for hele sin diskusjon om vitellogenin pseudogener hos mennesker er en artikkel av Brawand et al. 2008. Ann Gauger gjennomgikk dette artikkelen her og fant:
Venema sitering for hele sin diskusjon om vitellogenin pseudogener hos mennesker er en artikkel av Brawand et al. 2008. Ann Gauger gjennomgikk dette artikkelen her og fant:
Dette argumentet ble basert på foreliggende bevis fra det opprinnelige artikkelen av Brawand et al., Som er nevnt ovenfor, som rapporterte de opprinnelige tilpasninger av de synntiske vitellogeninregioner av kylling, pungrotte og menneske. (Synteni refererer til justeringen av DNA, typisk mellom arter, i samme rekkefølge og med identifiserbar sekvensidentitet.) Den artikkelen rapporterte at det var et vitellogenin-genrester i menneskelig DNA i regionen der den skulle være. Dessverre publiserte de svært lite data som grunnlag for deres krav, og jeg mente ikke at det underbygges det kravet.
Så Venema's sitering fra Brawand et al. 2008 støtter ikke hans påstander om at disse pseudogenene er til stede hos mennesker. Gauger gjorde senere en grundigere analyse enn Brawand et al. gjorde. Hun fant ut at det faktisk er "mer likhet mellom kyllinger og mennesker enn Brawand-artikkelen rapporterte." Men hun finner:
Til tross for den økte tilpasningen, er dette på grensen av det som kan oppdages som en match. Det faktum at ting varierer fra justering til justering indikerer at matchen er svak. Hvis det noen gang var et humant vitellogenin-gen, er det nesten ingenting igjen av det opprinnelige genet. Swamidass [en teistisk evolusjonistisk biolog] forteller meg at gitt den lange tiden som har gått siden vår siste vanlige forfedre med kyllinger, er dette å forvente. Ja jeg vet. Men det er en alternativ forklaring - muligheten for at et vitellogenin-gen aldri var der til å begynne med.
 Selv etter å ha tatt hensyn til den økte tilpasningen, bemerker Gauger at hun skrev sitt opprinnelige innlegg "for å kritisere Venema's påstand om sterkt bevis for et menneskelig vitellogenin pseudogen" og at "den kritikken står fortsatt. Hans illustrasjon overskrider mye graden av likhet." Venema stoler på de samme sitatene i Adam and the Genome som han gjorde i artikkelen kritisert av Ann Gauger. Hennes kritikk står også overfor hans argumenter i sin bok. Beviset i artikkelen han siterer er svært svak.
Selv etter å ha tatt hensyn til den økte tilpasningen, bemerker Gauger at hun skrev sitt opprinnelige innlegg "for å kritisere Venema's påstand om sterkt bevis for et menneskelig vitellogenin pseudogen" og at "den kritikken står fortsatt. Hans illustrasjon overskrider mye graden av likhet." Venema stoler på de samme sitatene i Adam and the Genome som han gjorde i artikkelen kritisert av Ann Gauger. Hennes kritikk står også overfor hans argumenter i sin bok. Beviset i artikkelen han siterer er svært svak.
Til slutt hevder Venema at felles avstamning er demonstrert fordi mønstrene til disse pseudogenene passer til et "nøstet hierarki" som er i samsvar med standard fylogeni for mennesker og store aper (sjimpanser, gorillaer og orangutanger). Det kan være sant - i dette spesielle tilfellet. Det er imidlertid mange genetiske data som ikke passer til standard fylogeni for høyere primater som Venema ikke snakker om.
For eksempel studerte en 2007-studie vår genomiske likhet med sjimpanser og andre primater og fant at store deler av vårt genom ikke passer inn i det standardiserte "nøstede hierarkiet" av menneske-ape-relasjoner:
"For ca 23% av genomet vårt, deler vi ingen umiddelbar genetisk forfedre med vår nærmeste levende slektning, sjimpansen. Dette omfatter gener og eksoner i samme grad som intergeniske regioner. Vi konkluderer med at ca. 1/3 av generene våre begynte å utvikle seg som menneskespesifikke linjer før differensieringen mellom mennesker, sjimpanser og gorillaer fant sted.
(Ingo Ebersberger, Petra Galgoczy, Stefan Taudien, Simone Taenzer, Matthias Platzer og Arndt von Haeseler, "Mapping Human Genetic Ancestry," Molecular Biology and Evolution , 24 (10): 2266-2276 (2007)
For detaljer, se her. Da, i 2012 da gorilla-genomet ble sekvensiert, ble det oppdaget at 30 prosent av gorilla-genomet motsiger den antatte evolusjonære fylogenien av mennesker og aper -her. Som en av forskerne setter det, "Vi kan ikke bare tillempe til et enkelt tre på gen-for-gen-basis."
Så, det er mye i vårt genom - store deler faktisk - som ikke passer til standard fylogeni. Det er ikke alltid slik at "identiske mutasjoner ... er tilstede før disse to linjene skilles" (s. 36), som Venema setter det. Noen ganger er de genetiske dataene i motsetning til det forventede mønsteret til et "nøstet hierarki." Så stort er dette problemet at hele 23 % av vårt genom ikke plasserer mennesker som nærmest relatert til sjimpanser, noe som står i kontrast til standard evolusjonært tre.
Samlet ser det ut til at genetiske data passer til standard human-ape fylogeni, bortsett fra når det ikke gjør det. Evolusjonære biologer har som alltid deres etterfaktorer forklaringer (her, ufullstendig sortering av avstamningslinjer). Poenget er at store mengder av genomdata varierer slående fra Venema's eksempel på noen pseudogener som passer til standard fylogeni. Hvis det faktum at noen få pseudogener passer til standardfylogenien, på noen måte gir bevis for felles forfedre, gir ikke det faktum at over 20 prosent av vårt genom ikke passer til standardfylogenien, bevis mot felles forfedre?
Tager: Ann Gauger; ENCODE; felles avstamning; genomet; kromatin.
Oversettelse og bilder ved Asbjørn E. Lund